Ŕâňîđű: Marco Dorigo, Member, IEEE,Vittorio Maniezzo and Alberto Colorni Čńňî÷íčę: IJ.10-SMC96.pdf
The Ant System: Optimization by a colony of cooperating agents
Marco Dorigo, Member, IEEE,Vittorio Maniezzo and Alberto Colorni
Introduction
In this paper we define a new general-purpose heuristic algorithm which can be used to solve different combinatorial optimization problems. The new heuristic has the following desirable characteristics:
It is versatile, in that it can be applied to similar versions of the same problem; for example,
there is a straightforward extension from the traveling salesman problem (TSP) to the
asymmetric traveling salesman problem (ATSP).
It is robust. It can be applied with only minimal changes to other combinatorial optimization
problems such as the quadratic assignment problem (QAP) and the job-shop scheduling
problem (JSP).
It's population based approach. This is interesting because it allows the exploitation of
positive feedback as a search mechanism, as explained later in the paper. It also makes the system amenable to parallel implementations (though this is not considered in this paper).
These desirable properties are counterbalanced by the fact that, for some applications, the
Ant System can be outperformed by more specialized algorithms. This is a problem shared by
other popular approaches like simulated annealing (SA), and tabu search (TS), with which we
compare the Ant System. Nevertheless, we believe that, as is the case with SA and TS, our
approach is meaningful in view of applications to problems which, although very similar to
well known and studied basic problems, present peculiarities which make the application of the
standard best-performing algorithm impossible. This is the case, for example, with the ATSP.
In the approach discussed in this paper we distribute the search activities over so-called
"ants," that is, agents with very simple basic capabilities which, to some extent, mimic the
behavior of real ants. In fact, research on the behavior of real ants has greatly inspired our
work (see [10], [11], [21]). One of the problems studied by ethologists was to understand how
almost blind animals like ants could manage to establish shortest route paths from their colony
to feeding sources and back. It was found that the medium used to communicate information
among individuals regarding paths, and used to decide where to go, consists of pheromone
trails. A moving ant lays some pheromone (in varying quantities) on the ground, thus marking
the path by a trail of this substance. While an isolated ant moves essentially at random, an ant
encountering a previously laid trail can detect it and decide with high probability to follow it,
thus reinforcing the trail with its own pheromone. The collective behavior that emerges is a
form of autocatalyticbehavior1 where the more the ants following a trail, the more attractive that
trail becomes for being followed. The process is thus characterized by a positive feedback loop,
where the probability with which an ant chooses a path increases with the number of ants that
previously chose the same path.
Consider for example the experimental setting shown in Fig. 1. There is a path along which
ants are walking (for example from food source A to the nest E, and vice versa, see Fig. 1a).
Suddenly an obstacle appears and the path is cut off. So at position B the ants walking from A
to E (or at position D those walking in the opposite direction) have to decide whether to turn
right or left (Fig. 1b). The choice is influenced by the intensity of the pheromone trails left by
preceding ants. A higher level of pheromone on the right path gives an ant a stronger stimulus
and thus a higher probability to turn right. The first ant reaching point B (or D) has the same
probability to turn right or left (as there was no previous pheromone on the two alternative
paths). Because path BCD is shorter than BHD, the first ant following it will reach D before the
first ant following path BHD (Fig. 1c). The result is that an ant returning from E to D will find
a stronger trail on path DCB, caused by the half of all the ants that by chance decided to
approach the obstacle via DCBA and by the already arrived ones coming via BCD: they will
therefore prefer (in probability) path DCB to path DHB. As a consequence, the number of ants
following path BCD per unit of time will be higher than the number of ants following BHD.
This causes the quantity of pheromone on the shorter path to grow faster than on the longer
one, and therefore the probability with which any single ant chooses the path to follow is
quickly biased towards the shorter one. The final result is that very quickly all ants will choose
the shorter path.
The algorithms that we are going to define in the next sections are models derived from the study of real ant colonies. Therefore we call our system Ant System(AS) and the algorithms we introduce ant algorithms. As we are not interested in simulation of ant colonies, but in the use of artificial ant colonies as an optimization tool, our system will have some major differences with a real (natural) one:
artificial ants will have some memory,
they will not be completely blind,
they will live in an environment where time is discrete.

Fig. 1. An example with real ants.
a) Ants follow a path between points A and E.
b) An obstacle is interposed; ants can choose to go around it following one of the two different paths with equal probability.
c) On the shorter path more pheromone is laid down.
Nevertheless, we believe that the ant colony metaphor can be useful to explain our model.
Consider the graph of Fig. 2a, which is a possible AS interpretation of the situation of Fig. 1b.
To fix the ideas, suppose that the distances between D and H, between B and H, and between
B and D-via C-are equal to 1, and let C be positioned half the way between D and B (see
Fig. 2a). Now let us consider what happens at regular discretized intervals of time: t=0, 1, 2
Suppose that 30 new ants come to B from A, and 30 to D from E at each time unit, that each ant walks at a speed of 1 per time unit, and that while walking an ant lays down at time t a pheromone trail of intensity 1, which, to make the example simpler, evaporates completely and instantaneously in the middle of the successive time interval (t+1, t+2).
At t=0 there is no trail yet, but 30 ants are in B and 30 in D. Their choice about which way to go is completely random. Therefore, on the average 15 ants from each node will go toward H and 15 toward C (Fig. 2b).
At t=1 the 30 new ants that come to B from A find a trail of intensity 15 on the path that leads
to H, laid by the 15 ants that went that way from B, and a trail of intensity 30 on the path to C,
obtained as the sum of the trail laid by the 15 ants that went that way from B and by the 15 ants
that reached B coming from D via C (Fig. 2c). The probability of choosing a path is therefore
biased, so that the expected number of ants going toward C will be the double of those going
toward H: 20 versus 10 respectively. The same is true for the new 30 ants in D which came
from E.
This process continues until all of the ants will eventually choose the shortest path.

Fig. 2. An example with artificial ants.
a) The initial graph with distances.
b) At time t=0 there is no trail on the graph edges; therefore, ants choose whether to turn right or left
with equal probability.
c) At time t=1 trail is stronger on shorter edges, which are therefore, in the average, preferred by ants.
The idea is that if at a given point an ant has to choose among different paths, those which were heavily chosen by preceding ants (that is, those with a high trail level) are chosen with higher probability. Furthermore high trail levels are synonymous with short paths.
The Ant System
In this section we introduce the AS. We decided to use the well-known traveling salesman
problem [26] as benchmark, in order to make the comparison with other heuristic approaches
easier [20]. Although the model definition is influenced by the problem structure, we will show
in Section VII that the same approach can be used to solve other optimization problems.
Given a set of n towns, the TSP can be stated as the problem of finding a minimal length
closed tour that visits each town once. We call dij the length of the path between towns i and j;
in the case of Euclidean TSP, dij is the Euclidean distance between i and j (i.e.,
 ). An instance of the TSP is given by a graph (N,E), where N is the
set of towns and E is the set of edges between towns (a fully connected graph in the Euclidean
TSP).
). An instance of the TSP is given by a graph (N,E), where N is the
set of towns and E is the set of edges between towns (a fully connected graph in the Euclidean
TSP).
Let  be the number of ants in town i at time t and let m
be the number of ants in town i at time t and let m
 be the total number of ants. Each ant is a simple agent with the following characteristics:
be the total number of ants. Each ant is a simple agent with the following characteristics:
it chooses the town to go to with a probability that is a function of the town distance and
of the amount of trail present on the connecting edge;
to force the ant to make legal tours, transitions to already visited towns are disallowed
until a tour is completed (this is controlled by a tabu list);
when it completes a tour, it lays a substance called trail on each edge (i,j) visited.
Let  be the intensity of trail on edge (i,j) at time t. Each ant at time t chooses the next town, where it will be at time t+1. Therefore, if we call an iteration of the AS algorithm the m moves carried out by the m ants in the interval (t, t+1), then every n iterations of the algorithm (which we call a cycle) each ant has completed a tour. At this point the trail intensity is updated according to the following formula
be the intensity of trail on edge (i,j) at time t. Each ant at time t chooses the next town, where it will be at time t+1. Therefore, if we call an iteration of the AS algorithm the m moves carried out by the m ants in the interval (t, t+1), then every n iterations of the algorithm (which we call a cycle) each ant has completed a tour. At this point the trail intensity is updated according to the following formula
 (1)
(1)
where
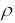 is a coefficient such that (1 - ) represents the evaporationof trail between time t and t+n,
is a coefficient such that (1 - ) represents the evaporationof trail between time t and t+n,
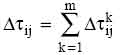 (2)
(2)
where  is the quantity per unit of length of trail substance (pheromone in real ants) laid on
edge (i,j) by the k-th ant between time t and t+n; it is given by
is the quantity per unit of length of trail substance (pheromone in real ants) laid on
edge (i,j) by the k-th ant between time t and t+n; it is given by
 (3)
(3)
where Q is a constant and Lk is the tour length of the k-th ant.
The coefficient must be set to a value < 1 to avoid unlimited accumulation of trail (see note
1). In our experiments, we set the intensity of trail at time ,  , to a small positive constant c.
, to a small positive constant c.
In order to satisfy the constraint that an ant visits all the n different towns, we associate with each ant a data structure called the tabu list2, that saves the towns already visited up to time t and forbids the ant to visit them again before n iterations (a tour) have been completed. When a tour is completed, the tabu list is used to compute the ant's current solution (i.e., the distance of the path followed by the ant). The tabu list is then emptied and the ant is free again to choose. We define tabuk the dynamically growing vector which contains the tabu list of the k-
th ant, tabuk the set obtained from the elements of tabuk , and tabuk (s) the s-th element of the list (i.e., the s-th town visited by the k-th ant in the current tour).
We call visibility  the quantity 1/dij. This quantity is not modified during the run of the AS, as opposed to the trail which instead changes according to the previous formula (1).
We define the transition probability from town i to town j for the k-th ant as
the quantity 1/dij. This quantity is not modified during the run of the AS, as opposed to the trail which instead changes according to the previous formula (1).
We define the transition probability from town i to town j for the k-th ant as

where allowedk = {N - tabuk} and where and are parameters that control the relative importance of trail versus visibility. Therefore the transition probability is a trade-off between visibility (which says that close towns should be chosen with high probability, thus implementing a greedy constructive heuristic) and trail intensity at time t (that says that if on edge (i,j) there has been a lot of traffic then it is highly desirable, thus implementing the autocatalytic process).
The algorithms
Given the definitions of the preceding section, the so-called ant-cyclealgorithm is simply stated
as follows. At time zero an initialization phase takes place during which ants are positioned on
different towns and initial values ij(0) for trail intensity are set on edges. The first element of
each ant's tabu list is set to be equal to its starting town. Thereafter every ant moves from town
i to town j choosing the town to move to with a probability that is a function (with parameters
and see formula (4)) of two desirability measures. The first, the trail ij(t), gives information
about how many ants in the past have chosen that same edge (i,j); the second, the visibility ij,
says that the closer a town the more desirable it is. Obviously, setting = 0, the trail level is no
longer considered, and a stochastic greedy algorithm with multiple starting points is obtained.
After n iterations all ants have completed a tour, and their tabu lists will be full; at this point
for each ant k the value of Lk is computed and the values kj are updated according to formula
(3). Also, the shortest path found by the ants (i.e., min Lk, k = 1, ..., m) is saved and all the
tabu lists are emptied. This process is iterated until the k tour counter reaches the maximum (user-defined) number of cycles NCMAX , or all ants make the same tour. We call this last case
because it denotes a situation in which the algorithm stops searching for
alternative solutions.
Formally the ant-cyclealgorithm is:
1.Initialize:
Set t:=0 {t is the time counter}
Set NC:=0 {NC is the cycles counter}
For every edge (i,j) set an initial value =c for trail intensity and = 0 Place the m ants on the n nodes
2. Set s:=1 {s is the tabu list index}
For k:=1 to m do
Place the starting town of the k-th ant in tabuk(s)
3. Repeat until tabu list is full {this step will be repeated (n-1) times}
Set s:=s+1
For k:=1 to m do
Choose the town j to move to, with probability pkj (t ) given by equation (4)
{at time t the k-th ant is on town i=tabuk (s-1)}
Move the k-th ant to the town j
Insert town j in tabuk (s)
4. For k:=1 to m do
Move the k-th ant from tabuk (n) to tabuk (1)
Compute the length Lk of the tour described by the k-th ant
Update the shortest tour found
For every edge (i,j)
For k:=1 to m do
 5. For every edge (i,j) compute ij(t+n) according to equation ij(t+n)= . ij(t)+ ij
Set t:=t+n
Set NC:=NC+1
For every edge (i,j) set ij:=0
6. If (NC < NCMAX) and (not stagnation behavior)
then
Empty all tabu lists
Goto step 2
else
Print shortest tour
Stop
5. For every edge (i,j) compute ij(t+n) according to equation ij(t+n)= . ij(t)+ ij
Set t:=t+n
Set NC:=NC+1
For every edge (i,j) set ij:=0
6. If (NC < NCMAX) and (not stagnation behavior)
then
Empty all tabu lists
Goto step 2
else
Print shortest tour
Stop
Ŕâňîđű: Marco Dorigo, Member, IEEE,Vittorio Maniezzo and Alberto Colorni Čńňî÷íčę: IJ.10-SMC96.pdf