Впервые проблемой поведения муравьев заинтересовались этологи, которые хотели понять как почти слепые животные, такие как муравьи, могли находить кратчайший путь от их гнезда к пище и обратно. Было выяснено, что муравьи общаются посредством следов феромона, отложенного на путях, которыми они следуют. Муравей откладывает феромон на землю, образуя след. Отдельно взятый муравей двигается случайно, тогда как муравей, который наткнулся на первоначально отложенный след, с большой вероятностью решит следовать этим путем, таким образом, откладывая новый слой феромона. Появление такой формы коллективного поведения называется автокаталитическим поведением - чем больше муравьев идут по следу, тем больше муравьев выберут этот путь.
Рассмотрим пример, показанный на рис.1. Здесь изображен путь, по которому прогуливаются муравьи (например от источника пищи А к гнезду Е, и наоборот рис 1а). Внезапно препятствие появляется и преграждает путь. Таким образом, в месте В муравьи, идущие от А к Е (или в месте D от Е к А) должны решить повернуть направо или налево рис.1b. На выбор влияет интенсивность феромонового следа, оставленного предшествующими муравьями. Поэтому большее количество феромона справа, стимулирует муравьев поворачивать направо. Первый муравей, достигший точки B (или D) с равной вероятность может повернуть как направо, так и налево (т.к. на альтернативных путях еще нет феромона). В следствие того, что путь BCD короче чем BHD, первый муравей идущий этим путем достигнет точки D быстрее муравья, шедшего путем BHD (рис. 1c). Таким образом, муравей возвращающийся из Е в D найдет более сильным след на пути DCB, появившийся из-за того, что половина муравьев решила обойти препятствие путем DCBA и некоторые достигли цели через BCD: в дальнейшем вероятно они предпочтут путь DCB пути DHB. Как следствие, количество муравьев последовавших путем BCD за единицу времени будет больше, чем количество на пути BHD. Это приводит к тому, что отложение феромона на коротком пути происходит быстрее, чем на длинном и вероятность выбора короткого пути муравьем также возрастает. В результате вскоре все муравьи выбирают короткий путь.

Рисунок 1 - Пример с настоящими муравьями
- a) Муравьи идут по пути между точками А и Е.
b) Появление препятствия; муравьи с равной вероятностью выбирают один из путей обхода препятствия
c) На коротком пути отложено большее количество феромона
Тем не менее, мы полагаем, что пример с муравьями может быть полезен для объяснения нашей модели. Рассмотрим граф на рисунке 2а, который является возможной интерпретацией с помощью муравьиной системы ситуации на рисунке 1b.
Предположим, что расстояние между D и H, между B и H, и между B и D- через C-равны 1, и пусть C располагается на полпути между D и B (смотри рис 2a. Теперь рассмотрим, что будет проистходить в интервалы времени t=0, 1, 2.
Предполагается, что 30 новых муравьев идут в точку В из точки А, и 30 в D из E в каждый момент времени, который каждый муравей проходит со скоростью 1 за каждый момент времени. Пока муравей идет, он откладывает в момент времени t феромон с интенсивностью 1, и , чтобы сделать пример проще, испаряется полностью и мгновенно во временной интервал (t+1, t+2). В момент времени t=0 следа еще нет, но 30 муравьев в точке В и 30 в D. Их выбор каким путем пойти полностью случаен. Вследствие этого, в среднем 15 муравьев из каждого узла будут двигаться по направлению к H, и 15 по направлению к C (рис. 2b) В момент времени t=1 30 новых муравьев, которые шли в В из А обнаружат след с интенсивностью 15 на пути, который ведет к Н, оставленный 15 муравьями, которые шли этим путем из точки В, и след с интенсивностью 30 на пути С, полученный как сумма следа отложенного 15 муравьями которые шли из В и 15, которые пришли в В из D через С (рис. 2c)
Этот процесс продолжается пока все муравьи случайным образом выберут короткий путь.
Рисунок 2 - Пример с искусственными муравьями
- a) Начальный граф с расстояниями
b) В момент времени t=0 нет никакого следа на ребрах графа; далее муравьи выбирают повернуть налево или направо с равной вероятностью
c) В момент времени t=1 след сильнее на коротком ребре, потому как он был предпочтительней для муравьев
Муравьиная система
В этой части представлена муравьиная система (AS). Было решено использовать известную задачу коммивояжера, чтобы сравнить с другими вероятностными методами.
Дано множество из n городов, задача коммивояжера представляет собой проблему нахождения самого короткого пути, при условии, что каждый город был посещен лишь раз. Обозначим dij - длина пути между городами i и j; в случае эвклидовой задачи коммивояжера dij - эвклидово расстояние между i и j; (т.е.Пусть ![]() количеств муравьев в городе i в момент времени t
количеств муравьев в городе i в момент времени t
![]() - общее количество муравьев. Kаждый муравей это простой агент со следующими характеристиками:
- общее количество муравьев. Kаждый муравей это простой агент со следующими характеристиками:
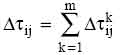 (2)
(2) (3)
(3)
 5. Для каждого ребра (i,j) вычисляется ij(t+n) в соответствии с формулой
5. Для каждого ребра (i,j) вычисляется ij(t+n) в соответствии с формулой