Точное крупнозернистое моделирование красных кровяных клеток
Игорь Пивкин и Джордж Эм Карниадакис
Отдел прикладной математики, Университет Брауна, Провиденс, Род-Айленд 02912, США
(Поступила в Редакцию 6 сентября 2007 года; опубликована 12 сентября 2008 г.)
Мы разрабатываем систематическую крупнозернистую процедуру для моделирования красных кровяных телец (эритроцитов) с использованием аргументов, основанных на теории среднего поля. Трехмерная модель мембраны эритроцита учитывает энергию изгиба, энергию в плоскости сдвига и ограничения фиксированной площади поверхности и фиксированной замкнутого объема. Крупнозернистая процедура носит общий характер, она может быть использована для произвольного уровня грубой зернистости и не используют каких-либо подгоночных параметров. Чувствительность крупнозернистой модели исследуется и его поведение проверяется на имеющихся экспериментальных данных в диссипативной динамике частиц, моделируя эритроциты в капиллярных и сдвиговых течениях.
Человеческий эритроцит имеет двояковыпуклую форму с диаметром около 8 мкм. Мембрана клетки состоит из двухслойного липида и прилагаемого цитоскелета. Цитоскелет состоит в основном из белков спектрина, которые образуют сеть, которая образована короткими нитями актина распределительных комплексов. Важность понимания механических свойств эритроцитов мотивируется рядом экспериментальных и теоретических исследований. На основе молекулярных численных моделей были разработаны численные методы и применены к моделированию эритроцитам. Уровень детализации мембраны эритроцитов описанных в этих исследованиях варьируется. Разработанная модель уже использовалась в работе и описывала РБК спектриновые сети с разрешением вплоть до отдельных комплексных переходов, но речь идет о 3х10000 степени свободы. Это было успешно проверено на экспериментальных данных механических реакций индивидуальных клеток, однако применение модели в процессе моделирования требует слишком много затрат и вычислений.
В этой статье мы разрабатываем систематическую крупнозернистую процедуру, которая позволяет уменьшить число степеней свободы в модели эритроцита на 2 порядка.
Вместе с крупнозернистой поточной моделью, метод диссипативной динамики частиц может привести к эффективному моделированию эритроцитов в микроциркуляции.
Модель мембраны состоит из совокупности точек, которые являются вершинами поверхности триангуляции красных кровяных клеток. Площадь треугольника сформированного
вершинами (l,m,n) задается х )/ 2. Длина ссылки соединяющих вершины m и n в триангуляции дает .
Свободная энергия Хельмхольца системы:
В плоскости свободный терм энергии:
включает в себя червеобразные цепи потенциальные для отдельных звеньев
где максимальная длина из ссылки и р является сохранение длины; параметр C
в гидростатическом упругой энергии.
Изгибная Энергия находится:
где k[bend] - cредний изгиб модуля, а Q0 и Qa - спонтанные и мгновенные углы
между двумя соседними треугольниками, соответственно. Известные выражения для общего объема и площади известны ранее. Силы получаются из , в то время как эволюция скорости и координат типичной точки n [1..N] описывается уравнениями движения Ньютона:
, где Vn - скорость и Fext внешняя сила, приложенная на точкy.
Модель использовалась ранее и составляла N=23867 точек, каждая из которых представляет переход комплекса в РБК спектриновой сети.Средняя длина связи L0 было 75 нм. Максимальная длина Lmax расширении было принято быть 3.17хL0 длина р=7.05 нм.
Средний изгиб модуля был kbend=200kBT и спонтанный угол Q=1 градус. Модели показали результаты, которые
хорошо согласуются с экспериментальными данными оптических пинцетов. (Мы будем относиться к этой модели, как идеальная модель и будем помечать буквой "f" далее в тексте.)
В серии экспериментов было показано, что эритроциты подлежат переходу сдвигового течения релаксации после восстановления двояковыпуклой формы, в которой краи ячейки всегда образованны той же частью мембраны. Эти результаты предполагают, что есть упругая энергии, запасенной в мембране компонентов, что местные компоненты мембраны без напряжения сохраняют двояковыпуклую свободную форму.
Учитывая состояние исходного материала для плоскостной упругой энергии выбрана двояковыпуклая модель.Средняя форма безударных двояковыпуклые РБК измерена экспериментально и состоит из множество точек с
координатами (х, у, z) в 3D-пространстве, удовлетворяющая следующему уравнению:
,где (R,c0,c1,c2)=(3,91 нм,0.0135805,1.001279,0.561381). Мы использовали следующую процедуру, чтобы получить начальные двояковыпуклые формы: Во-первых, точки равномерно распределены по аналитической форме и используются в качестве вершин поверхности триангуляции. Далее, движение точек ограничено двояковыпуклыми формами, в то время как их позиции обновляются в связи с релаксацией свободной энергии модели. Во время этого процесса, ссылки меняются местами, чтобы гарантировать, что для каждой пары соседних углов грани между треугольников короче диагонали соединяющих пары противоположных вершин. Конечное состояние получается после достаточно долгого времени уравновешивания. Мы разрабатываем крупнозернистую модели, используя меньше
количество точек (N <23 867), чтобы представить эритроцит. Равновесная длина связей зависит от числа используемых ччастиц(точек). Простое приближение, основанное на геометрическом аргументе среднего увеличения мембранной
области за каждую точку:
здесь далее получаются хорошие результаты, где Nf и Nc это числа точек в тонкой и крупнозернистого моделях, соответственно, и Lc0 является равновесной длиной связей в крупнозернистой моделе. Средний угол между парами
смежных треугольников увеличивается по мере использования крупнозернистых моделей. Поэтому, используя аналогичный геометрический аргумент, мы регулируем спонтанный угол:
Для настройки параметров в плоскости сдвига уравнения энергии мы используем аргумент среднего поля. Апроксимация среднего поля является аналитической. Для получения модуля сдвига клеточной мембраны используем:
Таблица 1 - Эффективные параметры модели на различных уровнях грубой зернистости при температуре T = 300 К.
Параметры описаны в тексте.
и модуль упругости сжатия области:
Заметим, что при фиксированном значении х0=L0/Lmax, сдвига и упругой области сжатия модулей из грубого
зернистой мембраны равны оригинальным модулям, если длина стойкости регулируется как:
Системы уравнений (4), (8), (9) и (12) дают нам модели с полным набором параметров при произвольном уровне грубой зернистости, до тех пор, пока число точек не будет достаточным для описания деформации ячейки. Параметры модели для различных значения грубой зернистости использовались ранее в тексте и перечислены в таблице 1.
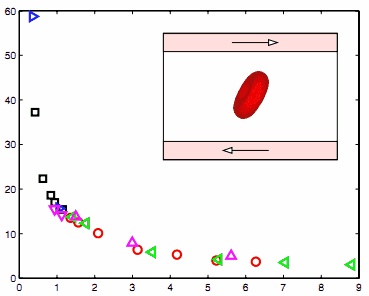
Рис. 1. Осевые и поперечные диаметры РБК относительно растягивающей силы. Оптический пинцет экспериментальных
данных показан символами. Результаты моделирования на различных уровнях грубой зернистости показаны линии. Синяя
линия: N 23867, красная линия: N 5000, зеленая линия N 500 и
пурпурный: N 100. Примечание: Стойкая длина чувствительности исследования для N 500. Синяя линия: р задан в соответствии с формулой(12). Пурпурная линии: р=7.5 нм. Зеленая и красная линии: р установлен на 10%
выше или ниже значения, даваемого формулой(12), соответственно.
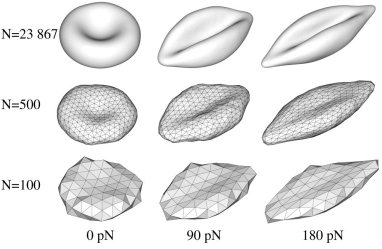
Рис. 2. Форма эволюции эритроцитов при различных силах растяжения (0, 90,
и 180 pN) предсказанные моделями на различных уровнях грубой зернистости. Верхний ряд: N 23 867 (поверхность триангуляции не показаны), средний ряд: N 500, нижний ряд: N 100.
Чтобы убедиться в грубой зернистости процедуры, мы впервые образовали множество клеточных растяжений и проводили эксперименты на различных уровнях грубой зернистости. Данные, используемые для исследования характеристики модели была взята из оптического пинцета в экспериментах [3]. Первоначально модель покоится с большим диаметром находясь в плоскости хz. Мы находим 5% от точек с координатами X крупнейших. Во время моделирования, мы применяем зависящих от времени силы Fext. Применяется к 5% пунктов с наименьшим х-координаты в состоянии покоя. Осевые и
поперечный диаметры вычисляются как Max(x)-Min(x). Результаты моделирования представлены на рис. 1 и
хорошо согласуются с экспериментальными данными. Для N 100 модель дает очень грубое округление поверхности клетки, как показано на рис. 2. Мы начали наблюдать отклонения продольного и поперечного диаметров, которые становятся более выраженные как у крупнозернистых моделей дальше (N 50, здесь не показана). Таким образом, мы делаем вывод, что N>100, необходимых для представления точности деформации РБК. Для оценки чувствительности результатов для грубой зернистой модели на параметры, выполняем дополнительное моделирование, где мы используем штраф настойчивости модели. Длина (то есть, положим р=7.5 нм) или назначать его на 10% выше, или ниже значения, даваемого формулой(12). В более поздних случаях мы наблюдали линейное отклонение продольного и поперечного диаметров, тогда как в первом случае модель дает ошибочный однородный результат, как показано на вставке на рис. 1 для N 500. Далее, мы применяем крупнозернистую модель для имитации движение клетки крови используя микроканальную диссипативную
динамику частиц, метод мезоскопии, который описывает кластеры молекул, движущихся вместе в Лагранжевом моде и имеют мягкие квадратичные потенциалы. Эритроцит моделируется как набор DPD N частиц. Эта модель является погруженным в жидкость DPD. Клетки-частицы взаимодействуют с частицами жидкости через DPD потенциалов и температуры системы
контролируется с помощью термостата DPD. Деформация и движения эритроцита рассчитываются на основе формулы (6),
, в которой внешние силы, FEXT происходит от взаимодействия с окружающей жидкостью DPD. Область течения представляет собой трубку, 45 нм в длину и 10нм в диаметре. В капиллярях такого диаметра, скорость крови, как правило, около 1 мм/с. Условие прилипания на жидкость-твердое интерфейс достигается за счет использования адаптивного управления с равномерным распределением плотности предписано в пристеночной области. Первоначально жидкость покоится и клетка крови помещается в середину канала. Мы применяем силу на тело осевого направления и включаем поток в трубе. Эритроцит деформирует под условия движения и после некоторого переходного периода
принимает форму парашюта, показанного на рис. 3, что обычно наблюдается в экспериментах. После того как
сила выключена поток замедляется и в конечном итоге DPD жидкости возвращается в состояние покоя, в то время как эритроцит восстанавливает свою равновесную двояковыпуклую форму. Результаты показаны на рис. 3, показано N 500, но аналогичные результаты получены для более изысканные модели (например, N 5000), если параметры
масштабируется надлежащим формулам. (4), (8), (9) и (12). Использование DPD, мы также моделировали движение
клетки в сдвиговом потоке.
Рис. 3. Последовательные снимки деформации крупнозернистого модели (N 500) от моделирования клетки в DPD в микроканальных потоках. Слева направо: (а) модель помещается в канал с покоящейся жидкости. (б), (в) деформации
Модель 0,008 и 0,016 секунды после тела движущей силой потока применяется. (г) форма модели в устойчивом поток. (е)
Модель восстанавливает свою равновесную двояковыпуклую форму за 0,2 секунды после выключения силы.
Рис. 4. Переход от встряски к покою
крупнозернистая (500 Н, 1000,5000, соответственно) клетка модель сдвигового течения в DPD моделировании.
Касательно напряжения, зависимого от перехода движения, может быть объяснен на основе упругого хранения энергии в мембране эритроцитов. В этой статье мы разработали крупнозернистую модель клетки крови и применили ее в симуляции DPD потока. Модель может быть использована с другими частицами основных методов, таких как
сглаженных частиц (SPH) или решетки Больцмана(ЛБМ), а также на сплошной основе.
Литература
R. P. Rand and A.C. Burton, Biophys. J. 4, 115 (1964).
T.M. Fischer, Biophys. J.86, 3304 (2004)
J. P. Mills, L. Qie, M. Dao, C. T. Lim, and S. Suresh, Mech. Chem. Biosys. 1, 169 (2004).
M. Abkarian, M. Faivre, and A. Viallat, Phys. Rev. Lett.98, 188302 (2007).
G. Tomaiuolo, V. Preziosi, M. Simeone, S. Guido, R.
Ciancia, V. Martinelli, C. Rinaldi, and B. Rotoli, Ann Ist
Super Sanita 43, 186 (2007).
P.B. Canham, J. Theor. Biol. 26, 61 (1970).
W. Helfrich, Z Naturforsch C 28, 639 (1973).
U. Seifert, K. Berndl, and R. Lipowsky, Phys. Rev. A 44,
1182 (1991).
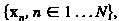 , которые являются вершинами поверхности триангуляции красных кровяных клеток. Площадь треугольника
, которые являются вершинами поверхности триангуляции красных кровяных клеток. Площадь треугольника 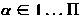 сформированного
вершинами (l,m,n) задается
сформированного
вершинами (l,m,n) задается 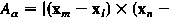 х )/ 2. Длина ссылки
х )/ 2. Длина ссылки 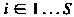 соединяющих вершины m и n в триангуляции дает
соединяющих вершины m и n в триангуляции дает 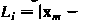 .
. 

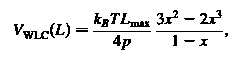
 максимальная длина из ссылки и р является сохранение длины; параметр C
в гидростатическом упругой энергии.
максимальная длина из ссылки и р является сохранение длины; параметр C
в гидростатическом упругой энергии.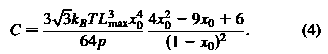
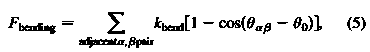
 , в то время как эволюция скорости и координат типичной точки n [1..N] описывается уравнениями движения Ньютона:
, в то время как эволюция скорости и координат типичной точки n [1..N] описывается уравнениями движения Ньютона: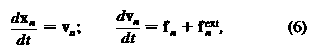



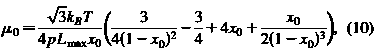

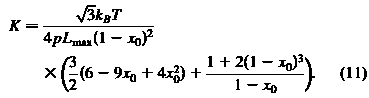
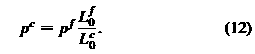


 к покою
к покою крупнозернистая (500 Н, 1000,5000, соответственно) клетка модель сдвигового течения в DPD моделировании.
крупнозернистая (500 Н, 1000,5000, соответственно) клетка модель сдвигового течения в DPD моделировании.