Изучение параметров флуоресценции хлорофилла в листьях древесных растений, растущих в условиях г. москвы
Д.И. Орехов, Ю.П. Козлов
Экологический факультет
Российский университет дружбы народов
Подольское шоссе, 8/5, Москва, Россия, 113093
Аннотация
В статье рассматриваются вопросы изучения основных изменений параметров флуоресценции фотосинтетического аппарата древесных пород (береза, тополь, можжевельник) под влиянием загрязнения в САО г. Москвы (микрорайон Западное Дегунино).
Город Москва характеризуется напряженной экологической обстановкой с сильной степенью угнетения древесных насаждений. Флуоресцентный метод фитоиндикации может помочь при выявлении ослабленных и больных пород деревьев в различных местообитаниях города. Метод импульсной флуорометрии, основанный на измерении флуоресценции хлорофилла, широко применяется в экологических исследованиях. Метод основан на том, что при изменении состояния фотосинтетического аппарата растений под воздействием поллютантов или неблагоприятной окружающей среды происходят изменения квантового выхода, вероятности тушения энергии электронного возбуждения молекул хлорофилла и скорости относительного электронного транспорта. Изменения в квантовом выходе дают нам представление об эффективности использования энергии света в фотосинтетических клетках растений.
Материалы и методы исследования. Объектами исследований служили высечки коры и листья (хвоя) деревьев, произрастающих в микрорайоне Западное Дегунино САО г. Москвы, по функциональному состоянию которых определялась их устойчивость к загрязнителям. Образцы коры и листьев деревьев брались 2 раза в месяц с октября 2009 г. по апрель 2010 г. Для экспериментов были отобраны модельные деревья. Деревья были примерно одинакового возраста и развития. Опытными считались деревья, произрастающие в местах сильного загрязнения атмосферы вблизи автодороги (Дмитровское шоссе). Для контроля выбирались деревья из относительно чистых мест обитания (пруд, школа), защищенных от действия газов. Все измерения проводились на одних и тех же ветках в нижней части на высоте 1,5–2 м от земли в утренние часы. Повторность измерений 10‑ кратная. Непосредственно перед началом анализа у побегов отделялся участок коры (0,5–1,5 см2) с одного и того же места по длине. Листья и побеги перед началом проведения измерений выдерживались в темноте около 10 мин. в увлажненном состоянии. Активность фотосинтетического аппарата листьев и феллодермы регистрировали по параметрам флуоресценции на импульсном портативном флуорометре PAM‑210 (Walz, Германия):
1) квантовый выход флуоресценции – Fv/Fm;
2) скорость относительного электронного транспорта;
3) коэффициенты фотохимического и нефотохимического тушения флуоресценции хлорофилла при разных интенсивностях света (от 0 до 786 мкЕ) определяли с использованием компьютерной программы PamVin.
Результаты и их обсуждение.Условия произрастания растений в разных районах города и в разных типах садово‑парковых насаждений неодинаковы. Так, в сомкнутых посадках деревья страдают меньше, чем при раздельном произрастании. На окраинах парковых массивов деревья повреждаются больше, чем в центральных участках. Наиболее неблагоприятные условия складываются для деревьев в уличных посадках и на территории промышленных предприятий. Несмотря на пространственную неоднородность городской среды, можно выделить ряд местообитаний от пригорода к центру, составляющих экологический профиль города, который характеризует нарастание комплекса городских влияний (в первую очередь ухудшение почвенных условий, загрязнения воздуха). Загрязнение воздуха на магистралях и улицах превышает допустимые для растений пределы, причем негативное воздействие загрязнения воздуха продолжается на протяжении всего года, а в летнее время оно усугубляется как за счет увеличения числа автомобилей, так и за счет поражения наиболее чувствительной части растений – листьев. Как правило, наиболее уязвимы для поражения деревья и кустарники, примыкающие к проезжей части магистралей и улиц. Именно на этих участках обнаружено наибольшее число ослабленных и усыхающих деревьев, причем из‑за влияния автомобилей снижается естественный иммунитет растений, и они становятся гораздо более чувствительными для различного рода заболеваний и вредителей. Факторы, связанные с автотранспортом, наиболее токсичны для растений из‑за повышенного содержания в воздухе окислов азота и серы, а в почвах – тяжелых металлов. Максимальные концентрации окислов азота (в 3–6 раз выше допустимого уровня) обнаружены на МКАД и крупных магистралях города. По мере снижения интенсивности транспортного движения загрязненность воздуха закономерно снижается, и этот фактор перестает быть для растений существенным. Это касается только районов, достаточно удаленных от крупных дорог и перекрестков, как правило, дворовых территорий в жилых кварталах. Вредное прямое воздействие газов на растения проявляется непосредственно на листовом аппарате. Косвенное влияние газов на микрофлору, почвенный поглощающий комплекс, корни растений осуществляется через почву. Кислые газы нарушают водный режим растений (снижается оводненность тканей, падает содержание связанной воды), приводят к закислению цитоплазмы, изменению работы транспортных систем. Вследствие повреждения мембран хлоропластов снижается интенсивность фотосинтеза, разрушается хлорофилл a и каротиноиды. Особенно ядовитым для фотосинтеза считается сернистый газ, который лучше растворяется в воде, чем углекислота. На пигментную систему неблагоприятно действует хлор, сернистый газ. Наиболее чувствительны к воздействию ферменты фотосинтеза. Интенсивность дыхания вначале повышается, а затем, по мере усиления повреждения и отсутствия поставки субстратов вследствие нарушения фотосинтеза, снижается. Инактивируется транспорт электронов и всех окислительно‑восстановительных ферментов. Нарушается фосфорный обмен, содержание общего фосфора падает. Следует отметить, что деревья способны до определенного уровня загрязнения вырабатывать механизмы, благодаря которым они избавляются от токсических веществ. Одним из таких механизмов является накопление токсинов в листьях с последующим их сбрасыванием. Повышенное содержание вредных веществ в атмосфере приводит к изреживанию верхнего яруса даже тех деревьев, которые относительно устойчивы к загрязнению. Это дает возможность свету проникать под полог крон и, таким образом, способствует кустарникам или травянистой растительности. Подобного рода перераспределение света и пространства позволяет частично компенсировать снижение количества древесной биомассы и, соответственно, оздоровить городскую атмосферу. Однако в городской среде под кронами деревьев обычно располагается не земля (исключение составляют разве что парки), на которой могло бы что‑то вырасти, а каменная или заасфальтированная поверхность.
Рассмотрим изменения фотосинтетических параметров флуоресценции у тополя, березы и можжевельника, связанные с устойчивостью деревьев и кустарников не только к загрязнению воздуха, но и к сезонным фазам вступления растений в покой и выхода из покоя. В октябре параметры фотосинтеза оставались еще высокими. Значения квантового выхода фотосинтеза тополя вблизи шоссе соответствуют 0,2, а вблизи школы 0,3–0,56. Значения электронного транспорта и угла наклона флуоресценции тоже почти в 3 раза варьируют с загрязненным районом. Хорошо видна разница в значениях фотохимического тушения 0,81–0,55 у школы и 0,35–0,37 – у дороги. Нефотохимическое тушение флуоресценции большую долю имеет вблизи шоссе. Это связано с большей сопротивляемостью растений у дороги к негативным факторам загрязнения. Ближе к холодному периоду начинают сглаживаться значения по квантовому выходу, что видно при сравнении можжевельника и березы – значения рядом с шоссе 0,29–0,31, рядом с прудом и школой соответственно 0,56–0,5. По‑видимому, у деревьев начинают приостанавливаться основные физиологические процессы и значения нивелируются (сглаживаются). В целом, активность пока сохраняется на том же уровне, что и у тополя в конце октября. Обратная картина складывается, когда наблюдаются значения угла наклона по электронному транспорту у можжевельника. Если у березы наблюдается заметное разграничение в районе школы и шоссе, то у можжевельника рядом с шоссе углы наклона несколько больше по значениям, хотя квантовый выход намного ниже, чем у растений пруда. Вероятней всего, во‑первых, близость автомобильной дороги заметно продлевает время вхождения в покой, тогда как в нормальных условиях (пруд) фотосинтез должен уже снижать свою активность заблаговременно. Во‑вторых, можжевельник является хвойной породой, следовательно, фотосинтез продолжается в течение всего года и в связи с невозможностью сброса листвы в осеннее время можжевельник вынужден искать альтернативные адаптивные пути механизма устойчивости к загрязнению, поэтому наблюдается увеличение скорости потока электронов в транспортной цепи. В январе и феврале у тополя очень глубокий покой и значения сильно снижены, вблизи шоссе активность почти нулевая, света мало, собственно, и хлорофилл практически выгоревший, он почти отсутствует и равен 0,01. Этот факт объясняет, почему в больших городах деревья зимой практически сухие. Все остальные виды – можжевельник и береза – практически также держатся у нуля. В данном случае февраль определяет минимальный уровень активности всех деревьев. В марте уже начинается выход из покоя тополей, растущих вблизи пруда, тогда как на шоссе они по‑прежнему в сильном покое. Квантовый выход тополей на пруду и на шоссе соответствует значениям 0,37–0,3–0,25 и 0,18–0,07. Квантовый выход фотосинтеза у берез, растущих рядом со школой и шоссе, соответственно 0,18–0,22 и 0,02–0,15. У берез отмечаются более индифферентные значения и у школы, и у шоссе, причем береза и тополь, растущие рядом с шоссе, раньше выходят из покоя, судя по значениям электронного транспорта. Это связано с высоким уровнем стресса, которому подвержены деревья, растущие у дороги, по сравнению с удаленными районами. Однако у березы более выражена тенденция к выходу из покоя вблизи шоссе. В апреле происходит закономерное изменение активности деревьев в зависимости от местоположения. По значениям квантового выхода и электронного транспорта хорошо видна зависимость влияния автотрассы на деревья (табл.). По квантовому выходу значения у тополя между шоссе и прудом составляли 0,2 и 0,32, у березы 0,15 и 0,34, а у можжевельника 0,13 и 0,27–0,32 соответственно. Также заметно возрастание давления стресса у тополя по значениям нефотохимического тушения, как и в октябре, рядом с шоссе происходит закрытие реакционных центров в ответ на загрязнение, у березы и можжевельника эти значения равны нулю, что свидетельствует о сильном подавлении рядом с шоссе и недостатком энергии для фотосинтеза возле пруда. В целом апрель характеризуется выравниванием фотосинтетических параметров, что связано с началом вегетационного периода, связанного с важнейшим процессом образования листовых пластин из почек.
Для всех пород общее сходство выражается в снижении фотосинтетической активности в марте–апреле. Это еще раз показывает, насколько энергоемок процесс цветения и начала созревания. Хорошо разграничено значение квантового выхода весной и осенью у всех трех пород. У древесных пород активность весь год прямолинейно зависит от значений температуры. Осенью активность держится на высоком уровне из‑за влияния высоких летних температур в предыдущем периоде. Весной, несмотря на плавное повышение температуры, растение, пока оно не зацветет и у него не распустятся листья, не достигает высоких значений фотосинтеза (рис. 1).
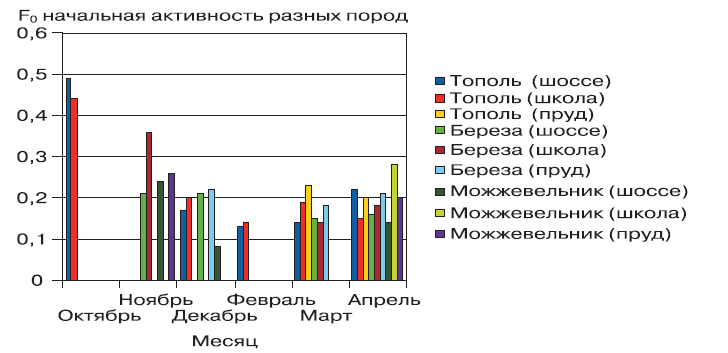
Рисунок 1 – Значения начальной активности (F0) фотосинтеза разных пород по месяцам
Минимальное содержание хлорофилла для тополя отмечалось в феврале. Начальная активность зимой небольшая, только после распускания листьев она начинает повышаться (рис. 2).
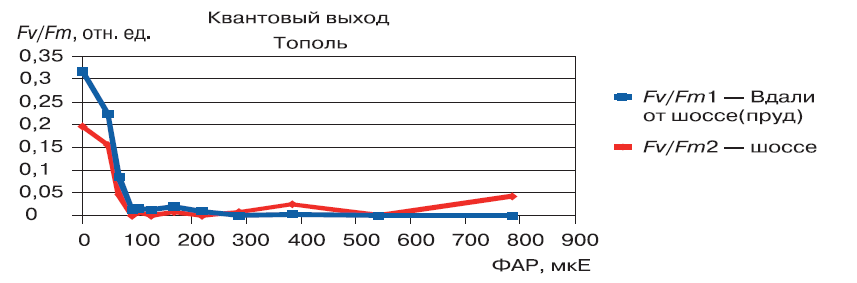
Рисунок 2 – Изменения параметра квантового выхода (Fv/Fm) фотосинтеза в зависимости от температуры (T) в период с октября 2009 по апрель 2010 г. (на примере тополя)
Коэффициент максимальной утилизации световой энергии α, рассчитанный по зависимости относительной скорости фотосинтетического транспорта электронов (rETR) от освещенности (апрель 2010 г.)
| Параметр | Тополь у шоссе | Тополь вдали от шоссе (пруд) | Береза у шоссе | Береза вдали от шоссе (пруд) | Можжевельник у шоссе | Можжевельник вдали от шоссе (пруд) |
| α | 0,0652 | 0,0935 | 0,0543 | 0,1022 | 0,0196 | 0,0522 |
Выводы.Выявлены сезонные изменения основных фотосинтезирующих параметров деревьев: квантового выхода фотосинтеза (Fv/Fm), относительного электронного транспорта (ETR), фотохимического тушения (qP), нефотохимического тушения (qN) на разном удалении от дороги. Как правило, деревья, растущие рядом с дорогой, подвержены большему стрессу, раньше выходят из покоя и длина их вегетационного сезона продлевается, в силу чего растения быстрее исчерпывают свои внутренние защитные резервы.
На степень ослабленности деревьев большое влияние оказывают почвенные условия. Степень рыхлости почвы, pH, наличие питательных веществ (азота, фосфора и калия) в достаточных количествах, тяжелых металлов, наличие (отсутствие) подземных коммуникаций, уплотненность и запечатанность почв – важнейшие факторы, влияющие на развитие деревьев.
Наиболее чувствительный период в жизни деревьев к загрязнению атмосферного воздуха связан с началом созревания и роста, приходящийся на март – апрель. В данный неблагоприятный период необходим особый тщательный уход за состоянием деревьев и особенно за состоянием почв.
По мере приближения к шоссе значения квантового выхода (Fv/Fm) уменьшаются, скорость относительного электронного транспорта (ETR) уменьшается, однако перед началом периода вегетации у березы и тополя, растущих вдоль шоссе, значения нивелируются по сравнению с удаленными от шоссе районами, угол наклона также снижается. Фотохимическое тушение (qP) до марта снижается, после чего увеличивается, нефотохимическое тушение (qN) возрастает в районе шоссе до момента вхождения в покой, после наступления покоя понижается (декабрь). Количество хлорофилла увеличивается пропорционально увеличению среднемесячных значений температуры и влажности.
Литература
1. Громов А.А., Щукин В.Б. Газоустойчивость растений. – Оренбург, 2002.
2. Фролов А.К. Окружающая среда крупного города и жизнь растений в нем. – СПб.:
Наука, 1998.
3. Чиркова Т.В. Физиологические основы устойчивости растений. – СПб.: Изд‑во Санкт‑
Петербургского ун‑та, 2002. – С. 21–122.
4. Bukhov N.G., Sridharan G., Egorova E.A., Carpentier R. Changes in polyphasic chlorophyll a
fluorescence induction curve upon inhibition of donor or acceptor side of photosystem II in
isolated thylakoids // Biochim. et Biophys. Acta. – 2004. – 1657. – P. 121–130.
5. Buschmann C., Langsdorf G., Lichtenthaler H.K. Imaging of the blue, green, and red fluorescence
emission of plants: an overview // Photosynthetica. – 2000. – Vol. 38. – № 4. –
P. 483–491.